Darwin also war der Meinung, dass die Natur keine Sprünge mache. Dementsprechend hatte seine Evolutionstheorie einen gradualistischen Charakter: Die Entwicklung geht in kleinen Schritten vor sich und ihre Akkumulation über die Zeit führt unter der Regulierung durch die natürliche Auslese zum Spektrum der Arten, wie wir es kennen. Diese Vorstellung ist im wesentlichen auch in die moderne synthetische Theorie übergegangen. Dabei wird aber oft eine Unterscheidung zwischen Mikro- und Makroevolution gemacht: Der erstere Begriff bezieht sich auf Vorgänge der Speziation, d.h. der Bildung neuer Arten aufgrund eines schon bestehenden Bauplanes. Dabei ist es allerdings möglich, dass es z.B. bei der pionierhaften Besiedlung eines neuen Lebensraumes zu einer gemessen am evolutionären Zeitmassstab explosiven Entwicklung kommen kann; man spricht in diesem Fall von einer "adaptiven
Radiation".45Vgl. Siewing 1982: 117.
Mit dem Begriff der Makroevolution soll demgegenüber die Entstehung neuer Organisationstypen wie z.B. der Chordaten, Mollusken oder Arthropoden angesprochen sein. Kausal wird diese Unterscheidung mit einer dazu parallelen Differenzierung der Mutationen in Mikro- und Makromutationen erklärt. Siewing findet diese Gegenüberstellung "unglücklich", wohl deshalb, weil es bei der orthodoxen Theorie keinen Unterschied zwischen einer Mikro- und einer Makroevolution geben
kann:46Siehe Siewing 1982: 354-355.
Alle Veränderungen gehen ausnahmslos auf die allmähliche Summierung von kleinen Schritten zurück; diese erfolgt einfach langsamer oder schneller. Dazu Siewing:
Wir kennen heute keine solchen Faktoren, durch die schlagartig ganze Organisationstypen entstehen können. Die Evolution ist demnach eine kontinuierliche Folge von Artbildungsvorgängen, von Artumbildungen, von Umbildungen der Merkmale im Sinne einer Anpassung; so sind sicher [sic!] auch die grossen Baupläne, die Organisationstypen entstanden: schrittweise, durch additive Typogenese.47Siewing 1982: 117.
Für viele aber ist es völlig undenkbar, dass ein schrittweiser Mechanismus von Mutation, Rekombination, Selektion und Isolation als Erklärung ausreicht, und zwar nicht nur für den komplexen Fall der Bildung neuer Arten oder gar neuer Baupläne, sondern schon für den relativ einfachen Fall der Entwicklung eines neuen Organs bei einer bestimmten Art. Koestler schildert dazu das Beispiel des Grossen
Pandas.48Siehe Koestler 1981: 128.
Dieser besitzt an seinen Vorderextremitäten einen sechsten Finger, der für die Handhabung der Bambussprossen, die seine hauptsächliche Nahrung darstellen, sehr nützlich ist. Damit er aber ein funktionstüchtiges Organ ist, braucht es Knochen, Muskeln und Nerven. Die Wahrscheinlichkeit, dass in der gleichen Population unabhängig voneinander passende, d.h. einander sinnvoll ergänzende, die Knochen, Muskeln und Nerven betreffende Mutationen stattfinden würden, dürfte wohl ausserordentlich klein sein. Hinsichtlich solcher undenkbarer Kombinationen meint denn auch Koestler: "The doctrine that their coming together was due to a series of blind coincidences is an affront not only to commonsense but to the basic principles of scientific
explanation."49Koestler 1981: 129.
Es muss also, damit die Entstehung eines solchen Organs überhaupt möglich wird, ein irgendwie koordinierendes Prinzip dazu treten.
Umso schwieriger dürfte es sein, sich vorstellen zu können, wie z.B. ein Übergang vom Wasser- zum Landleben oder umgekehrt ohne Zwischenformen zustande kommen kann. Nehmen wir das Beispiel des
Fischsauriers:50Nach Rieppel 1988.
Dieser verfügt bei seinem ersten Auftreten über ein bereits fertig gestaltetes Paddel und es existieren keine Zwischenformen, die die sukzessive Umbildung der fünfstrahligen Extremität der landlebenden Vorfahren belegen würden. Es kann natürlich nicht bestritten werden, dass das Paddel für einen Meeresbewohner einen selektiven Vorteil bietet, aber es ist nicht einzusehen, dass es einen schrittweisen Anpassungsprozess gegeben haben könnte, bei dem auch jedes Zwischenstadium überlebensfähig war. In Fällen wie diesem wird der Begriff des Funktionswandels zur Erklärung
herangezogen.51Siehe dazu ausführlich Josef H. Reichholf 1992: speziell145 ff.
Entwicklungen wie die gerade genannte sind nicht durch eine Ansammlung von Einzelmerkmalsveränderungen zustande gekommen, sondern durch die funktionelle Abwandlung ganzer Organe. Z.B. wird aus einer Schwimmflosse ein Arm, aus einem Arm ein Flügel, bei Beibehaltung der Grundbausteine, die diese Organe zusammensetzen (vgl. Abb.3). Das heisst dann aber, dass der genetische Apparat in der Lage sein muss, nicht nur für Einzelmerkmale, sondern für ganze zusammenhängende Merkmalskomplexe zu kodieren. Und wie es dann eine sprunghafte Änderung zu einer wiederum sinnvollen neuen Kodierung kommen kann, bleibt weiterhin eine Frage.
Das Auftreten von Lücken im Fossilienbeleg hatte Darwin damit zu erklären versucht, dass die Aufzeichnung der Evolutionsgeschichte in den Gesteinen einfach unvollständig
sei.52Siehe Leakey und Lewin 1995: 19-20.
Er nahm an, die betreffenden Lebewesen hätten entweder aus irgendwelchen Gründen keine Fossilien gebildet, oder wenn es Fossilien gab, wären diese durch spätere geologische Aktivitäten zerstört worden, oder auch, die fraglichen Fossilien würden vielleicht nur auf alten, heute versunkenen Kontinenten vorkommen. Heute aber beginnen sich die Stimmen zu mehren, die diese Lücken in einem positiven Sinne als Anzeichen dafür betrachten, dass es im Fortgang der Evolution in der Tat sprunghafte Ereignisse gegeben hat. So haben 1972 Nils Eldredge und Stephen Jay Gould die Theorie der
punctuated equilibria, also der Gleichgewichte, die gelegentlich unterbrochen werden,
aufgestellt.53Siehe Nils Eldredge und Stephen Jay Gould 1972, hier dargestellt nach Rieppel 1987 und 1988. Vgl. auch Gould 1986: 259-260.
In der Folge reden wir vom Punktualismus im Gegensatz zum Gradualismus. Danach bleibt eine Art während der Dauer ihrer Existenz unverändert, zumindest mit Bezug auf ihre sie kennzeichnenden Eigenschaften. Diese Unveränderlichkeit hängt mit einer genetisch bedingten Kanalisation der Embryonalentwicklung zusammen, die die Art in bestimmte Entwicklungsbahnen zwingt. Damit wird der wachsende Organismus vor den Auswirkungen von schädlichen Punktmutationen oder auch von Umwelteinflüssen geschützt.
Damit ein evolutiver Sprung zustande kommt, ist also eine vorgängige "genetische Revolution" nötig, welche die Wirkung der Entwicklungszwänge lockert oder aufhebt. Evolution ergibt sich damit aus der Divergenz von embryonalen Entwicklungsbahnen, und es ist dann folgerichtig, dass Zwischenstufen in der Form von ausgewachsenen Organismen fehlen. Die kreative Rolle verschiebt sich von der Ebene der natürlichen Selektion zu der der Erbanlagen und der embryonalen Entwicklung. Diese kann als Prozess gesehen werden, bei dem kreative Potenzen der Neubildung einschränkenden Entwicklungszwängen gegenüberstehen. Wie das Genom vom einen zu einem anderen sinnvollen Plan für diese Entwicklung umschalten kann, ist aber damit natürlich auch nicht geklärt. Augros und Stanciu vermuten
folgendes:54Vgl. Augros und Stanciu 1987: 182.
Das im Moment überflüssige Material, das in jedem Genom vorhanden ist, entwickelt langsam einen neuen, vorläufig verborgenen Bauplan.
Regulationsgene55Regulationsgene sind solche Gene, die ganze Sequenzen von Strukturgenen ein- oder ausschalten können. Die Strukturgene produzieren die einzelnen Proteine.
sorgen dann dafür, dass dieser Plan in einem bestimmten Zeitpunkt aktiviert wird. Heisst dies, dass dann das neue Produkt zum vorneherein schon sinnvoll ist? Oder kann die Sache, wie Rieppel meint, grundsätzlich auch schief laufen, womit der Selektion immer noch eine Rolle hinsichtlich der Beurteilung des Ganzen zugewiesen
wäre?56Vgl. Rieppel 1987.
Koestler denkt, dass es bei den durch die auf der Erde gegebenen natürlichen Bedingungen hinsichtlich der biologischen Baupläne eine begrenzte Anzahl von möglichen Themen gibt: "The conclusion which emerges from all this is that there must be unitary laws underlying evolutionary variety, permitting unlimited variations on a limited number of
themes."57Koestler 1981: 147.
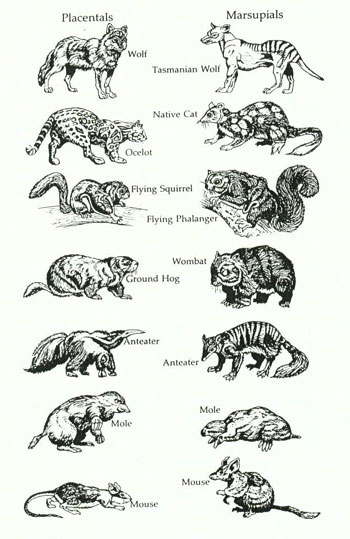
Er bringt dazu das Beispiel der "Doppelgänger", nämlich die Existenz einer verblüffenden Ähnlichkeit von verschiedenen Arten (z.B. Maulwurf, Ameisenbär, Maus, Flughörnchen, Wolf) der zwei Typen von Säugetieren, der Plazentatiere einerseits und der Beuteltiere andererseits, obschon sie auf voneinander getrennten evolutiven Pfaden entstanden sind (siehe Abbildung 4). Die orthodoxe Theorie muss diesen Umstand als eine "adaptive Ähnlichkeit" erklären, bewirkt durch den Druck der natürlichen Selektion bei entsprechend ähnlichen Umweltbedingungen. Das ist aber unsinnig, denn die Übereinstimmung der vergleichbaren Arten ist derart verblüffend, dass die entsprechenden Umwelten praktisch identisch hätten sein müssen, was nicht der Fall
ist.58Siehe Koestler 1981: 143.