Systematik |
Sie erfolgt nach abgestufter Ähnlichkeit. Eine kausale Erklärung ist nur durch die Annahme einer gemeinsamen, historischen, also stammesgeschichtlichen Herkunft möglich, vorausgesetzt, es handelt sich bei den Ähnlichkeiten um homologe. |
Vergleichende Anatomie |
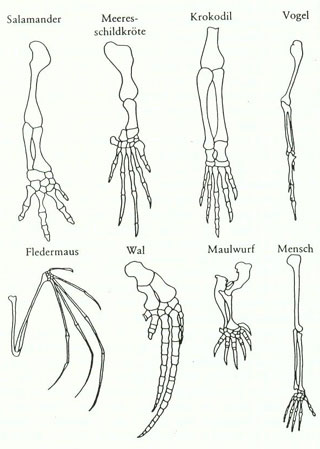
Beispiel für homologe Organe: Bei allen vierfüssigen Wirbeltieren haben die Vorderextremitäten immer den gleichen Bauplan, unabhängig davon, an welche Funktion (Laufen, Graben, Fliegen, Schwimmen) diese Extremitäten angepasst sind (vgl. Abbildung 3). Die Übereinstimmung lässt sich also nicht auf eine gemeinsame Funktion zurückführen, sondern sie kann durch die Annahme einer gemeinsamen, stammesgeschichtlichen Herkunft erklärt werden. |
Paläontologie |
Die fossil überlieferten Organismen können die Veränderungen im Laufe der Erdgeschichte darlegen. |
Embryologie |
Bereits anfangs des 19. Jh. lieferte die embryologische Forschung Zeugnisse für die Evolutionstheorie. Beispiel: Bei Hühnerembryonen treten Kiemenspalten und dazugehörige Blutgefässe auf, obwohl beim erwachsenen Hühnchen Kiemenspalten nicht vorhanden sind. Dies erklärt sich aus der stammesgeschichtlichen Vergangenheit: Es gibt eine übereinstimmende Embryonalentwicklung bei Hühnchen, Fischen und Amphibien. |
Tiergeographie |
Aus ihrem Bereich schöpften bereits Darwin und Wallace ihr Erfahrungsmaterial. Beispiel: Veränderung der Meisenarten bei ihrer Verbreitung von Zentraleuropa nach Osten. Es bilden sich verschiedene Rassen aus. |
Haustiere |
Darwin versuchte, seine Selektionstheorie mit Untersuchungen an Haustieren zu untermauern. Allerdings ist die Mannigfaltigkeit immer eine bloss intraspezifische. Beispiel: Unterschied zwischen der Widerristhöhe beim Bernhardiner (65-80 cm) und dem Zwergpinscher (26 cm). Mit anderen Worten: Aus keiner Züchtung ist bisher eine neue Art entstanden. |
Verhaltensforschung |
Diese hat Ergebnisse vorgelegt, die nur unter dem Gesichtspunkt der Evolution gedeutet werden können. |
Rudimentäre Organe (evtl. auch Verhaltensmuster) |
Es handelt sich um Merkmale, die ihre ursprüngliche Funktion verloren haben. Beispiel: Funktionsloser Beckengürtel bei Walen durch die Reduktion der Extremitäten. Er bleibt aber in reduzierter Form erhalten. |